




Kveitekornet innanfrå: anatomi, maling og kva dette tyder for bakaren
Når du opnar ein mjølsekk og tek ei neve mjøl, held du i hendene resultatet av to ting samstundes: millionar av år med botanisk evolusjon — og eit teknologisk inngrep som heiter maling. Mjøl er ikkje berre «innmaten» i kornet.
For nokre år sidan ville vi gjere reisene våre litt meir varierte med ferie i Danmark. Eg leita etter eit slott der ein kunne leige rom og kvile utanom turisttida. Eg byrja tidleg, og då eg fann eit, skreiv eg til eigarane. Men det synte seg at dei ikkje planla så langt fram i tid, og dei var enno ikkje sikre på korleis dei ville leige ut det vesle ætteslottet sitt. Mellom anna reklame på nettsida såg eg då eit tilbod om opphald på ei mølle. Ei mølle er betre enn eit slott, tenkte eg, og vi reiste til ei lita bygd med ei enorm mølle, 22 meter høg og med ei historie på nær et hundre og åtti år. Den venlege og gjestfrie huseigaren synte seg å vere son til møllaren og tilsynsmann for dette underlege bygget.

Møllaren synte oss tynne trepinnar eller trespiler med namn skrivne på. Når dei faste kundane kom, visste møllaren kva slags maling dei trong; han stakk denne trækile inn mellom møllesteinane, og den store konstruksjonen lydde han og mol kornet til mjøl av den kvaliteten som trongst. Det er heilt klårt at for å forstå mjøl, må ein forstå kornet. Møllesteinane tek lag for lag av kornet, og kjem gradvis inn til kjernen. Difor må ein, for å forstå korleis korn vert mala, først forstå korleis det er bygd opp.
Kornet som system: tre vevssoner
Kveitekornet — botanisk kalla karyopse — er ikkje eit frø, men ei frukt. Frukthinna (perikarp) har vakse tett saman med frøskalet og dannar ei udeleleg einskap. Hos spelt er det grunnleggjande det same: også her er kornet botanisk ei karyopse, ikkje eit frø. Perikarpen er på same måten samanvaksen med frøskalet. Skilnaden ligg ikkje i sjølve frukttypen, men i morfologien til karyopsen og i forholdet hennar til dei ytre blomsterdekkja: hos spelt ligg karyopsen tett innelukka i agnehinnene, og dette påverkar teknologien for handsaming av kornet sterkt — det må først avskallast. Dette førehandssteget verkar mellom anna inn på prisen på speltmjøl.
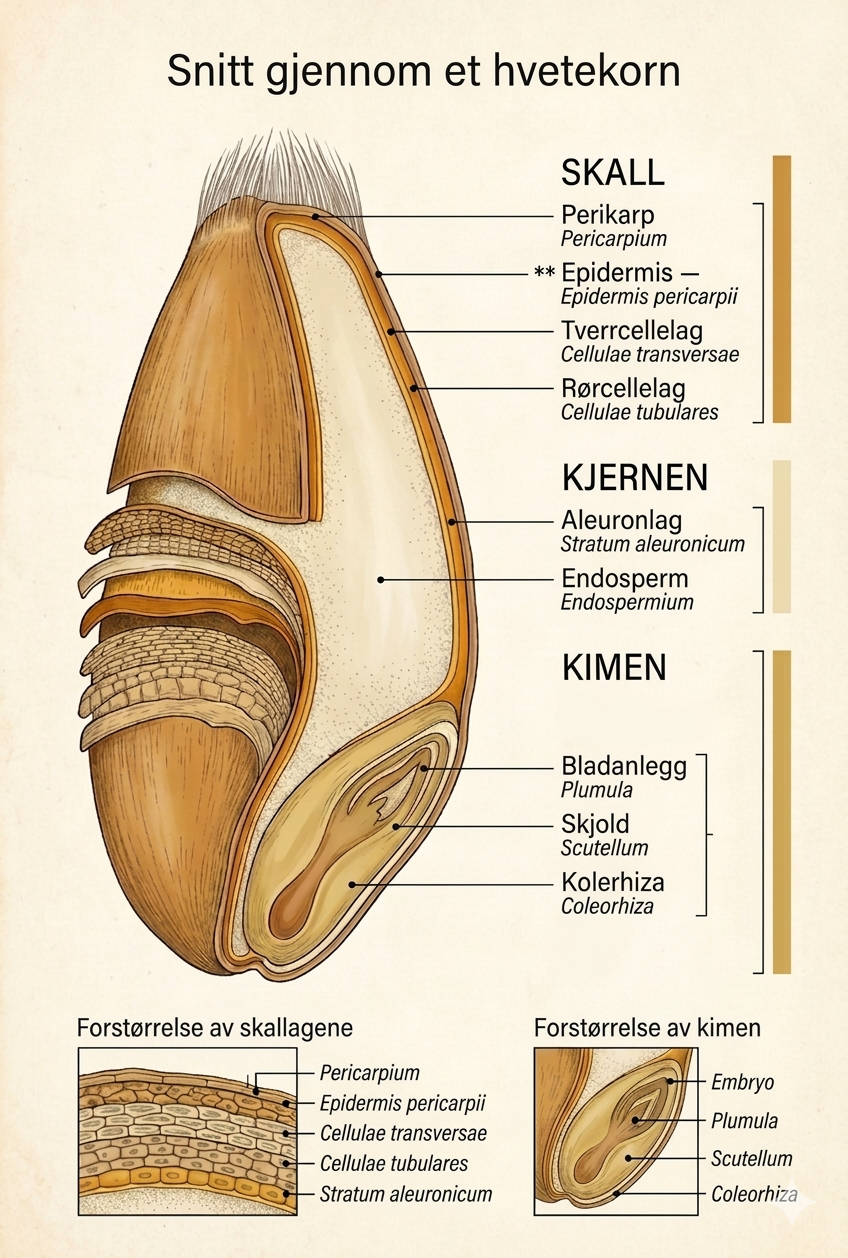
På figuren er kornet delt inn i tre store område: SKALL, KJERNEN og KIMEN. For bakaren og møllaren er dette ei nyttig og oversiktleg framstilling. Botanisk er røynda meir samansett. Særleg gjeld dette yttersona: figuren viser perikarpen og nokre av dei viktigaste cellelaga i perikarpen, men skil ikkje ut testa og nucellarlaget som eigne strukturar. I den faglege teksten nedanfor vert desse difor omtala uttrykkeleg der det er nødvendig, sjølv om dei ikkje er namna særskilt på skjemateikninga.
Såleis består kveitekornet av tre hovudområde med ulikt biologisk opphav, ulik kjemisk samansetjing og ulik åtferd under maling:
- dei ytre morveva — på figuren samla under SKALL
- endospermet — på figuren hovuddelen av KJERNEN
- kimen — på figuren KIMEN
1. Dei ytre laga: perikarp, testa og nucellarlag — «kli» i teknologisk tyding
Dei ytre laga i kornet kjem frå morplanta og utgjer det som i mølleteknisk språk vanlegvis vert omtala som kli. Desse laga står for om lag 14–16 prosent av kornmassen; her ligg det meste av kostfiberet, ligninet og antioksidantane. Botanisk er dette fleire heilt ulike vev, men under maling oppfører dei seg langt på veg som ei funksjonell einskap. Her ligg ei grunnleggjande skilnad mellom biologisk og mølleteknisk tenking: for møllaren og bakaren vert dette i praksis éin perifer fraksjon; for botanikaren er det fleire vev med ulik opphavshistorie og ulik funksjon.
På figuren er berre delar av denne yttersona viste eksplisitt, nemleg Perikarp, Epidermis pericarpii, Cellulae transversae og Cellulae tubulares. Dette svarar til sentrale delar av frukthinna, men ikkje til heile den botaniske periferien. Det er difor viktig å lese figuren som ei pedagogisk forenkling, ikkje som ei fullstendig histologisk oppløysing av alle ytre lag.
Perikarp
Perikarpen (frukthinna) er eit samansett lag som omfattar ytre epidermis, hypodermis, parenkymatøs mesokarp og indre epidermis. Alle desse cellene er døde når kornet vert hausta, med strukturelle, vedaktige cellevægger. Den ytre epidermis er kutikularisert — han har ei voksaktig overflate som avgrensar vassutveksling med omgjevnadene.
Under utviklinga av kornet var dei indre cellene i mesokarpen — særleg tverrceller og rørceller — fotosyntetisk aktive. Ja, det går faktisk føre seg fotosyntese i kornet òg. Seinare vert desse cellene lyserte, det vil seie at dei bryt seg ned ved sjølvstyrt nedbryting og etterlet luftrom som frigjer plass for det ekspanderande endospermet.
På figuren svarar Tverrcellelag / Cellulae transversae og Rørcellelag / Cellulae tubulares til nett desse karakteristiske delane av perikarpen.
Testa
Testa (frøskalet) er eit tynt samanpressa lag med pigment — særleg i raude sortar — fenoliske sambindingar og eit suberinisert lag. Saman med nucellarlaget dannar dette ein vassavvisande og delvis vasstett barriere mot patogen og uttørking.
Dette laget er ikkje merkt særskilt på figuren. På den pedagogiske teikninga er det i praksis innlemma i det samla omgrepet SKALL, sjølv om det botanisk ikkje er det same som perikarp.
Nucellarlaget
Nucellarlaget er ei einaste rad samanpressa, tome celler mellom testa og aleuronlaget. Det har eit særmerkt dobbelt kutikulært lag på begge sider, noko som gjer det til ein særs selektiv barriere mellom det maternelle leiingssystemet og det avleidde endospermet.
Heller ikkje dette laget er teikna ut som eit eige namn på figuren. Det må ein vere medviten om når ein les teksten opp mot skjemaet: overgangen frå SKALL til KJERNEN er i røynda meir samansett enn figuren gjev inntrykk av.
Kondisjonering — det avgjerande steget før maling
Her byrjar det som kvar bakar og møllar bør vite. Dersom ein byrjar å male tørt korn direkte, vil dei sterkt vedaktige og sprø ytterlaga smuldre opp til støv, kome inn i mjølet og straks auke oskeinnhaldet, forverre fargen og svekkje bakeeigenskapane. Nett difor er kondisjonering — oppfukting og kviletid før maling — det første og viktigaste steget i moderne mølleteknologi. Kornet vert fukta og får kvile i fleire timar, til ytterlaga vert elastiske og byrjar å losne i større flak i staden for å verte til pulver.
Ei nyare og stadig meir utbreidd tilleggsoperasjon er debranning, det vil seie varsam mekanisk avtaking av dei ytre laga ved friksjon alt før hovudmalinga. Dette reduserer innhaldet av bakteriar og mykotoksin kraftig og aukar utbytet av reint kvitt mjøl. Moderne industrimøller kombinerer ofte kondisjonering og debranning som standardprosedyre, i motsetnad til den gamle mølla vi budde på i Danmark.
Ved debranning går kornet vanlegvis gjennom ei maskin der overflata vert handsama abrasivt, ved friksjon, stundom òg i kombinasjon med slagpåverknad. Kornet vert altså på eit vis lett avskrapa, utan at sjølve kjernen vert fullstendig øydelagd.
Dette er ikkje det same som vanleg reinsing av korn. Vanleg reinsing fjernar det som ligg utanpå kornet: stein, støv, framandlekamar, metallpartiklar eller ugrasfrø. Debranning fjernar derimot ein del av sjølve kornet — dei perifere veva. Det er ein grunnleggjande skilnad.
Ikkje sjeldan vert denne prosessen forveksla med avskalling. Omgrepa kan stundom liggje nær kvarandre, men er ikkje alltid identiske, fordi avskalling oftast viser til ei grovare fjerning av skal eller agnehinner. Hos spelt, til dømes, er avskalling eit eige førehandssteg som kjem før debranning i det heile kan verte aktuelt.
Kva gjev dette bakaren? Om debranning er gjort før maling, vil mjølet vanlegvis verte lysare, reinare i smaken, med mindre innhald av skalpartiklar og meir føreseielege reologiske eigenskapar. Samstundes vert ein del stoff som er konsentrerte i dei perifere laga, fjerna: mineral, kostfiber, ein del enzym og ulike bioaktive komponentar. Dette er alltid ei avveging:
betre teknologisk åtferd og kvitare mjøl ↔ mindre anatomisk heilskap i kornet
I samband med dette kjem eg alltid til å tenkje på noko anna. Som kjent fastsette kyrkja alt i gammal tid at brødet som skal tene som Lammet i den guddomlege liturgien, skal vere baka av det beste mjølet. I eldre tid tydde «det beste» sjølvsagt alltid sikta mjøl, fint male, kvitt og utan ureiningar. Men om reglane vart vedtekne i dag, når vi veit så mykje om næringsverdien i ulike mjøltypar, trur eg dei heilage fedrane ville ha påbode at prosforer skulle bakast av sammale mjøl, som er rikare på mineral og nyttige stoff, sidan nett eit slikt mjøl etter denne målestokken utan tvil ville vere det beste.
Vidare skal vi tale om aleuronlaget.
2. Aleuronlaget: levande barriere og næringsbombe
På figuren ligg Aleuronlag / Stratum aleuronicum innanfor SKALL og er plassert som den ytre delen av KJERNEN. Dette er pedagogisk godt grunngjeve, men her må det gjerast ei presis botanisk merknad: aleuronlaget er botanisk ein del av endospermet, ikkje ein del av skallet. Figuren gjev i så måte ei teknologisk og visuelt klår framstilling, ikkje ei full botanisk klassifisering.
Aleuronlaget er det ytste levande laget i endospermet, eitt cellelag tjukt, og ligg rett innanfor nucellarlaget. Dette er den einaste levande delen av det modne kornet utanfor kimen. Cellene er blokkforma, med karakteristisk tjukke doble cellevægger rike på arabinoxylanar og β-glukanar. Kjernane er bevarte — cellene lever.
Biokjemisk er aleuronlaget eit konsentrert lager av lipid, proteinlekamar med fytin og niacin, og hydrolytiske enzym, mellom dei amylasar og lipasar, som vert aktiverte under spiring. I ernæringsmessig meining er dette eitt av dei rikaste laga i heile kornet.
Aleuronlaget og maling — paradokset
Sjølv om aleuronlaget botanisk høyrer til endospermet, hamnar det ved vanleg valsemaling systematisk i klifraksjonen. Årsaka er mekanisk: nucellarlaget og aleuronlaget er så sterkt bundne til kvarandre at dei i praksis ikkje let seg skilje utan svært spesialiserte inngrep. Det doble kutikulære laget i nucellarepidermis og dei tjukke cellevægger i aleuronlaget grip inn i kvarandre på ein slik måte at ingen normal malingsintensitet skil dei reint.
For ernæringa er dette eit monaleg tap. Aleuronlaget er ein hovudkonsentrasjon av B-vitamin, mineral som fosfor, kalium, magnesium, jern og mangan, og ulike antioksidantar i kveitekornet. Alt dette går i stor grad ut av den kvite mjølfraksjonen. Difor er heilt kvitt mjøl ernæringsmessig utarma samanlikna med meir fullstendige fraksjonar. Det er ikkje bokstaveleg tomt, men det er sterkt redusert i biologisk og ernæringsmessig kompleksitet.
Det finst òg ei motside. Det same aleuronlaget er rikt på feitt og aktive lipasar. Nett difor harsknar sammale mjøl mykje raskare enn kvitt mjøl: feittet oksiderer, og lipasane held fram med å verke. Å bake med ferskt sammale mjøl er ikkje berre eit ideologisk val. Det er den einaste måten å få fullt utbytte av eigenskapane til aleuronlaget på. Dersom du lagrar sammale mjøl lenge, må du vere merksam på at det kan harskne, og då kan sjølv god baketeknikk ikkje lenger redde smaken.
3. Stivelsesendospermet: gradientar av styrke og smak
Endospermet utgjer om lag 80–84 prosent av kornmassen og er hovudmålet for møllaren. På figuren er dette vist som den store lyse hovuddelen av KJERNEN, merkt Endosperm / Endospermium, med aleuronlaget som randsona utanfor. Denne framstillinga er nyttig, men ein må hugse at endospermet ikkje er ei einsarta masse. Det er eit romleg differensiert system med minst tre funksjonelt ulike soner — og det er nett dette som gjer mølleteknologien til noko meir enn berre «å male opp».
Subaleuronlaget
Mellom aleuronlaget og det sentrale endospermet ligg subaleuronlaget, ei overgangssone på to til tre cellelag. Dette laget er ikkje merkt særskilt på figuren; på skjemateikninga er det i praksis innlemma i den samla framstillinga av KJERNEN. Fagleg er det likevel avgjerande å skilje det ut.
Cellene her har dei tjukkaste cellevægger i heile stivelsesendospermet, minst stivelsesinnhald og det høgaste proteininnhaldet. Det er registrert opp til om lag 54 prosent protein i subaleuronrik mjølfraksjon innanfor mjøl som samla berre held 12,5 prosent protein. Glutenproteina her har ein særmerkt samansetjing: laget er rikt på ω-gliadinar og spesifikke høgmolekylære undereiningar av glutenin.

Cellene i det sentrale endospermet er større, rikare på stivelse og har tynnare cellevægger. Proteininnhaldet er lågare i prosent enn i subaleuronlaget, men dette tyder ikkje automatisk dårlegare bakeeigenskapar. Tvert om ligg det her eit av dei viktigaste paradoksa i moderne mjølkunnskap.
Glutenparadokset — det viktigaste bakaren må vite
Her kjem eit punkt som bryt med den enkle førestellinga om at «meir protein alltid gjev betre brød».
Sjølv om dei perifere subaleuroncellene inneheld meir protein, er glutenkvaliteten for brødbaking ofte høgare i dei sentrale cellene i endospermet. I den sentrale sona er delen av dei høgmolekylære gluteninundereiningane større, og det er nett desse som i høg grad står for styrken i deigen, elastisiteten hennar og evna til å halde på gass.
Det er difor mogleg at mjøl frå meir sentrale endospermfraksjonar gjev det mest volumrike og stabile brødet, trass i lågare samla proteininnhald. Mjøl frå perifere fraksjonar, nærare subaleuron og aleuron, inneheld ofte meir strekkbare gliadinar og gjev ei deig som er mjukare, meir utflytande og vanskelegare å forme.
Konklusjonen er viktig: straumar frå dei tidlege maleomgangane kan vere rike på totalt protein, men likevel gje svak og klissete deig. Straumar frå dei seinare omgangane, der det sentrale endospermet dominerer, kan gje sterk deig og stabil gjæring.
Stivelseskorn: to klassar, to åtferder
Stivelsen i kveite finst i to hovudklassar av granulat: store A-granulat, som vert danna tidleg i utviklinga av kornet, og små B-granulat, som vert danna seinare. B-granulata har mykje større spesifikk overflate og tek opp meir vatn per gram. Mjøl med høg del B-granulat krev difor meir vatn i deigen. I tillegg gelatiniserer dei to klassane noko ulikt under baking, noko som påverkar strukturen i krummen.
Endospermhardleik — genet i arbeid
Eitt av dei viktigaste einskildgena for både møllaren og bakaren er Ha-lokuset på kromosom 5D. Hos mjuke kveitesortar produserer dette genet puroindolinprotein, som legg seg på overflata av stivelseskorna og verkar som smørjing mellom protein og stivelse. Ved maling bryt mjukt korn saman langs cellegrensene; stivelseskorna kjem ut relativt heile, og mjølet vert fint og silkeaktig, med låg grad av stivelsesskade. Slike sortar er ideelle til kjeks, vaflar og fin søtbakst.
Hos harde sortar manglar puroindolina eller fungerer ikkje. Protein og stivelse er då langt sterkare bundne til kvarandre. Kornet bryt ikkje primært saman langs cellegrensene, men gjennom sjølve cellene, og stivelseskorna vert skadde.
Stivelsesskade er ikkje nødvendigvis ein feil. Skadde granulat tek opp langt meir vatn, er meir tilgjengelege for amylasar og støttar lengre og djupare gjæring. Hard kveite gjev difor eit mjøl som kan vere framifrå til surdeig og lang autolyse, men som krev mykje meir presis styring av vassmengda.
4. Fureområdet og kimen: teknologiske utfordringar
På figuren er KIMEN vist som eit eige område og forstørra nedst til høgre. Der finn vi Bladanlegg / Plumula, Skjold / Scutellum og Kolerhiza / Coleorhiza. Dette er i samsvar med den grunnleggjande botaniske oppbygginga av kimen. Men før vi kjem til sjølve kimen, må vi stanse ved eit anna område som figuren ikkje framhevar særskilt: den langsgåande bukfura.
Fureområdet
Fureområdet er den djupe bukfura langs heile kornet, der den maternelle leiingsstrukturen låg og forsynte det veksande endospermet med sukker og aminosyrer. For møllaren skaper dette to konkrete vanskar.
Den første er mekanisk: restar av kli set seg fast i sjølve fura og let seg ikkje fjerne fullstendig med børstar. Sjølv på moderne møller med eigne børstemaskiner blir noko att i fura, særleg hos sortar med djup eller ujamn fold. Nett difor vert utmjølingsgraden aldri 100 prosent: fura held att både restar av perifere vev og ein del endosperm.
Den andre er hygienisk: fura er eit vern for mikroorganismar og for sporar knytte til mykotoksin, som ikkje let seg fjerne ved rein overflatereinsing. Dette er ei av hovudårsakene til at debranning har vorte så viktig i moderne mølleteknologi: ved å ta bort perifere lag og delar av furekantane reduserer ein den mikrobiologiske og toksikologiske belastninga på mjølet markant.
Kimen
Kimen med skjoldet utgjer berre om lag 2,5–3 prosent av kornmassen, men er eit konsentrat av feitt av høg kvalitet, protein og vitamin. Han er òg ei levande og metabolsk aktiv eining.
Ved maling vert kimen systematisk skild frå og teken ut — ikkje fordi han er uviktig, men fordi lipasane i han er særs aktive og ville redusere lagringstida til mjølet kraftig. Kveitekim vert difor ofte seld som eiga næringsrik vare, men i mjøl som skal lagrast lenge, er han eit problem meir enn ein fordel.
Frå botanikk til mølle: korleis kornet faktisk vert mala
Når dei to hovudfaktorane som styrer maling er klåre — lagstrukturen i kornet og hardleiken i endospermet — kan vi leggje dei saman og følgje heile malegangen.
Stega i valsemaling
Den moderne mølla er ikkje éi maskin, men ei rekkje påfølgjande operasjonar. Først kjem kondisjonering og eventuelt debranning. Deretter byrjar sjølve malinga.
Dei første drageomgangane opnar kornet ved hjelp av rifla valsar som roterer med ulik fart. Målet er å bryte opp kornet slik at dei ytre laga kan losne, utan at endospermet straks vert redusert til fint mjøl. Dei lignifiserte og etter kondisjonering elastiske ytterlaga bryt då reint og gjev større klistykke som lettare kan skiljast frå.
Det kritiske brotet skjer ved grensa mellom nucellarlaget og aleuronlaget. Heftinga her er så sterk at vanleg maling ikkje kan skilje dei. Aleuronlaget går difor med klifraksjonen, og saman med det forsvinn mykje av vitamin-, mineral- og antioksidantinnhaldet frå den kvite mjølfraksjonen.
Dei påfølgjande reduksjonsomgangane er retta inn mot finmaling av det frigjorde endospermet. Her viser hardleiken seg att: hardt korn krev fleire omgangar og meir energi, gjev meir stivelsesskade og ei meir karakteristisk kornete mjølstruktur. Mjukt korn gjev eit finare og meir silkeaktig mjøl med lågare grad av stivelsesskade.
Malestraumar og proteingradient
Sidan endospermet ikkje er einsarta, er heller ikkje malestraumane det. Mjøl frå dei tidlege omgangane, der perifere delar av endospermet vert tekne ut, er ofte rikare på totalt protein, men gjev ikkje nødvendigvis betre deig. Mjøl frå seinare omgangar, der det sentrale endospermet dominerer, kan vere meir stivelsesrikt, men likevel gje sterkare og meir elastisk deig.
Profesjonelle møller nyttar dette med fullt medvit: dei blandar straumar for å oppnå eit bestemt proteininnhald og bestemte bakeeigenskapar, eller dei held dei frå kvarandre og sel dei til ulike industrielle føremål. For bakaren tyder dette noko svært viktig: to typar mjøl med same deklarerte proteininnhald kan oppføre seg grunnleggjande ulikt i deigen, alt etter kva fraksjonar dei er sette saman av.
På gamle vindmøller, slik eg sjølv såg i Sillerup, kunne ein òg skilje mjøl etter kvalitet, men mykje grovare enn i moderne industrimøller. Først vart kornet mala på møllesteinar. Då fekk ein ikkje eit ferdig, einsarta mjøl, men ei blanding av finare mjølstoff, grovare partiklar, mellomgrove gryn og skalrestar. Deretter vart blandinga sikta. Alt etter kor fine siktene var, skilde ein ut fint kvitt mjøl, grovare uttak, gryn og kli.
Ein del av dei grovare mellomprodukta vart ofte mala om att og sikta på nytt for å få ut endå noko meir fint mjøl. Såleis hadde også den gamle møllaren ikkje berre éin type mjøl, men fleire ulike uttak frå det same kornet. Desse vart likevel ikkje skilde etter indre anatomiske soner i endospermet, slik moderne møller kan gjere, men etter meir praktiske teikn: finleik, reinleik, farge og bakeåtferd.
Den gamle møllaren blanda altså ikkje «indre anatomiske straumar frå kornet», men delar av malingsproduktet som skilde seg i finleik og reinleik. For bakaren tyder dette noko heilt enkelt, men viktig: mjøl med same namn kunne vere merkbart ulikt frå møllar til møllar.
Vassopptak: tre faktorar samstundes
Mjølet si evne til å ta opp vatn er resultatet av minst tre faktorar på ein gong:
- forholdet mellom A- og B-granulat av stivelse
- graden av stivelsesskade under maling
- proteininnhaldet og proteinfordelinga
Desse verkar saman og kan ikkje lesast isolert frå kvarandre. Eit «sterkt» mjøl med høgt proteininnhald frå mjuk kveite kan krevje mindre vatn enn eit tilsynelatande «svakare» mjøl frå hard kveite med høg grad av stivelsesskade — sjølv når proteinprosenten på papiret er den same.
Kva hamnar kvar: oppsummerande tabell
Her må det gjerast ei lita, men viktig presisering opp mot figuren. Figuren grupperer kornet i SKALL, KJERNEN og KIMEN. Tabellen nedanfor går eit steg lenger og bryt dette ned på eit meir presist anatomisk og mølleteknisk nivå.
| Botanisk struktur | Fraksjon ved maling | Merknad i lys av figuren |
|---|---|---|
| Perikarp | Kli | På figuren vist som del av SKALL |
| Testa + nucellarlag | Kli | Botanisk viktige, men ikkje særskilt namna på figuren; funksjonelt høyrer dei til overgangen mellom SKALL og KJERNEN |
| Aleuronlag | Kli i praksis | På figuren vist i KJERNEN; botanisk korrekt, men mølleteknisk følgjer det klifraksjonen |
| Subaleuronlag | Delvis mjøl, delvis perifere fraksjonar | Ikkje merkt særskilt på figuren; ligg innanfor KJERNEN |
| Sentralt stivelsesendosperm | Mjøl | Svarar til hovuddelen av Endosperm / Endospermium på figuren |
| Skjold + kim | Kimfraksjon | Svarar til KIMEN på figuren, med scutellum, plumula og coleorhiza |
Terminologisk etterord
Den vedvarande kløfta mellom botanisk og mølleteknisk språk er ikkje berre eit akademisk spørsmål. Når ein produsent skriv «endosperm» på emballasjen, meiner han ikkje eit eintydig homogent stoff, men eit uttak frå ein romleg ueinsarta fleirlagsstruktur. Når ein talar om «kli», meiner ein ei blanding av botanisk ulike vev — inkludert slikt som botanikken i strikt tyding reknar til endospermet. Og når ein seier «sammale mjøl», seier ein framleis ikkje alt: alt etter finleiken på malinga, graden av debranning, innblandinga av aleuronrike fraksjonar og forholdet mellom kjerne og periferi kan slikt mjøl oppføre seg svært ulikt i deigen.
Å bake med forståing krev ikkje laboratorium. Men det hjelper bakaren til å lese mjølopplysningar med eit kritisk blikk, forstå kvifor sammale mjøl oppfører seg annleis enn sikta mjøl, og kvifor det ikkje finst eitt eintydig svar på spørsmålet: kva er godt mjøl?
Svaret er alltid det same: godt til kva.
Sjå no absolutt dei to videoane som eg omsorgsfullt har førebudd for dykk ved hjelp av kunstig intelligens. Den første er på engelsk, og den andre er på norsk:





